



Ji bo pîşesaziya tutunê toza enzîma asîd protease ya pola xwarinê peyda dike

Danasîna hilberê
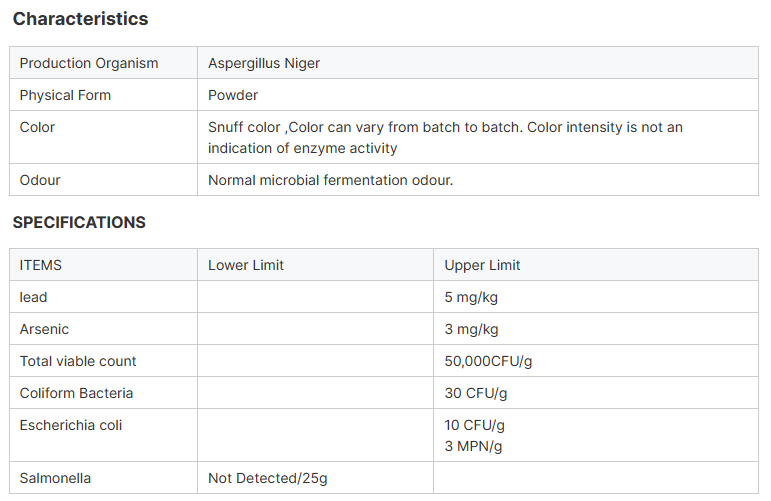
Acid protease celebek spehî ya aspergillus Nîjerê ye ku di nav fermentasyona kûr a şil, ultrafiltration û pêvajoyên din ên paqijkirî de ye, ew dikare di şert û mercên PH-ya nizm de be, bi çalakiya qutkirina hundurîn û derveyî ve ji bo hîdrolîzkirina proteînê di nav peptîd û asîdên amînî yên piçûk de, dawiya birrîna bijare aromatîkî ye an bermahiyên asîda amînî ya hîdrofobîk di navbera girêdana peptîdê de.


Karkirin
1.Zêdekirina asîdê proteazê ji bo hilweşandina proteîna pelên tûtinê dikare kalîteya şewitandina titûnê kêm bike, tama tûj, hêrs û tama tal kêm bike, û dereceya pelên tûtinê baştir bike.
2.Ew dikare bêhna tûtinê bi bandor dewlemend bike, tevna cixarekêşanê baştir bike, û bîhnfirehiya xwerû ya kok û gaza cûrbecûr kêm bike, da ku guheztina aroma çêtir be, û dikare germahiya dûmanê hevrêz bike, tama kokê kêm bike.
3.Pêkhatina kîmyewî ya navxweyî ya pelên tûtinê lihevhatîtir e û kalîteya hestiyar a pelên tûtinê çêtir dibe.
Rêbaza Serlêdanê
1. Doza ku tê pêşniyar kirin 50-300 gr enzîm ji bo her tonek madeya xav e. Pelên titûnê bi dûman bikin û bikin pelên;
2. Li gorî vê mîqdara sepandinê, rêjeyeke diyarkirî ya çareseriya amadekirina enzîmê hate pîvandin û bi alavên xwarinê yên ku ji xwe hatine çêkirin, li ser pelên titûna ezmûnî yeksan hate rijandin. Pelên titûnê ji bo hîdrolîza enzîmatîk di bin şert û mercên ceribandinê yên diyarkirî de ketin jûreyek germahî û nemtiyek domdar.
3.Piştî dermankirinê, pelên tûtinê di 120℃ de hatin neçalak kirin, perçe kirin û danîn aliyekî. Ji ber cûdahiya qada serîlêdanê û pêkhateya madeya xam û parametreyên pêvajoyê yên her kargehê, divê moda lêzêdekirina rastîn û mîqdara zêdekirina vê hilberê bi ceribandinê were destnîşankirin. .
Embarkirinî
Pakêt: 25 kg/drum; 1,125 kg / tembûr.
Hilanîn: Li cîhek zuwa û sar were girtin û ji tîrêja tavê dûr bisekinin.
Temenê rafê: 12 meh li cîhek hişk û sar.
Berhemên Têkildar:
Fabrîkaya Newgreen jî Enzîmên jêrîn peyda dike:
| Bromelain pola xwarinê | Bromelain ≥ 100,000 u/g |
| Protease alkaline pola xwarinê | Proteaza alkaline ≥ 200,000 u/g |
| Papain pola xwarinê | Papain ≥ 100,000 u/g |
| Laccase pola xwarinê | Laccase ≥ 10,000 u/L |
| Tîpa APRL ya proteaza asîdê ya pola xwarinê | Acid protease ≥ 150,000 u/g |
| Cellobiase pola xwarinê | Cellobiase ≥1000 u/ml |
| Enzîma dextranê asta xwarinê | Enzîma Dextran ≥ 25,000 u/ml |
| Lipase pola xwarinê | Lîpaz ≥ 100,000 u/g |
| Proteaza bêalî ya pola xwarinê | Proteaza bêalî ≥ 50,000 u/g |
| glutamine transaminase-pola xwarinê | Glutamine transaminase≥1000 u/g |
| Pectin lyase pola xwarinê | Pektîn lîaz ≥600 u/ml |
| Pectînaza pola xwarinê (avî 60K) | Pektînaz ≥ 60,000 u/ml |
| Katalaza pola xwarinê | Katalaz ≥ 400,000 u/ml |
| Glucose oxidase pola xwarinê | Glucose oxidase ≥ 10,000 u/g |
| alpha-amylase pola xwarinê (li hemberî germên bilind berxwedêr) | Germahiya bilind α-amylase ≥ 150,000 u/ml |
| alpha-amylase pola xwarinê (germahiya navîn) type AAL | Germahiya navîn alpha-amylase ≥3000 u/ml |
| Decarboxylase alpha-acetyllactate-pola xwarinê | α-acetyllactate decarboxylase ≥2000u/ml |
| β-amylase-pola xwarinê (avî 700,000) | β-amylase ≥ 700,000 u/ml |
| Tîpa β-glucanase BGS ya pola xwarinê | β-glucanase ≥ 140,000 u/g |
| Proteaza pola xwarinê (cureya endo-birrîn) | Proteaz (cureyê jêkirî) ≥25u/ml |
| Xylanase XYS-a pola xwarinê | Xylanase ≥ 280,000 u/g |
| Xylanase pola xwarinê (asîd 60K) | Xylanase ≥ 60,000 u/g |
| Tîpa GAL a glukoz amîlaz a asta xwarinê | Enzîma sakkarîker≥260,000 u/ml |
| Pullulanase pola xwarinê (liquid 2000) | Pululanase ≥2000 u/ml |
| Cellulaza pola xwarinê | CMC≥ 11,000 u/g |
| Cellulaza pola xwarinê (pêkhateya tevahî 5000) | CMC≥5000 u/g |
| Proteaza alkaline ya pola xwarinê (cûreya konsantrekirî ya çalakiya bilind) | Çalakiya proteaza alkaline ≥ 450,000 u/g |
| Glîkoz amylase pola xwarinê (100,000 zexm) | Çalakiya glukoza amylase ≥ 100,000 u/g |
| Protease asîdê pola xwarinê (50,000 zexm) | Çalakiya asîd protease ≥ 50,000 u/g |
| Proteaza bêalî ya pola xwarinê (cûreya konsantrekirî ya çalakiya bilind) | Çalakiya proteazê ya bêalî ≥ 110,000 u/g |
jîngeha fabrîkayê

pakêt & teslîmkirina


neqlîye











